|
||
DMSO Background Literature |
|||||||||||
|
L. Santos and P.G. Tipping Centre for Inflammatory Diseases • Monash University • Department of Medicine • Monash Medical Centre • Clayton • Victoria, Australia
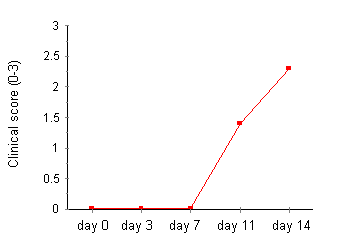
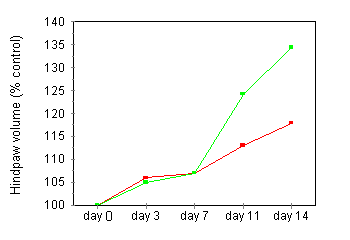
Development of Adjuvant Arthritis Polyarthritis was observed in all untreated rats immunized with MT in oil by day 14. The earliest onset of disease was observed at day 9. Fifty per cent of rats had evidence of arthritis by day 11 (clinical score 1.4 ± 0.3) and 100% by day 14 (2.3 ± 0.3; Fig.1). The hindpaw volume in control (oil injected) rats increased progressively over the observation period consistent with normal growth. Rats injected with MT in oil demonstrated significant hindpaw swelling on day 11 (normal 113.0 ± 1.4% day 1 volume, diseased 124.2 ± 5.2% day 0 volume, P < 0.05) and day 14 (normal 117.9 ± 3.4% day 0 volume, diseased 134.4 ± 8.5% day 0 volume, P < 0.05; Fig.1). Synovial lavage cells obtained from arthritic rats on day 14 contained 11.76 ± 1.7 x 105 cells/four hindlimb joints. This is significantly higher than cell numbers from normal rats (0.2 ± 0.1 x 105 cells/four hindlimb joints, P < 0.0001).
Phenotypic Characteristics of Synovial Cells of Arthritic Rats Synovial lavage cells (74.6 ± 3.0%) were
neutrophils as judged by nuclear morphology, T cells
expressing the
Phenotypic characteristic of cells harvested from arthritic joints.
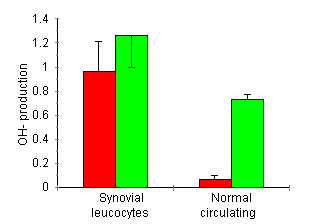
Hydroxyl radical production was measured with and without stimulation with PMA (Fig. 2). Synovial lavage cells from arthritic rats spontaneously produced OH· (0.96 ± 0.28 OH· units/h per 105 cells, P < 0.05 compared with normal leucocyte production). Stimulation with 10 mg PMA, resulted in a non-significant increase in OH· production (1.26 ± 0.29 OH· units/h per 105 cells). In contrast, spontaneous production normal circulating leucocytes was absent (0.07 ± 0.03 OH· units/h per l05) and increased significantly after PMA stimulation (0.73 ± 0.03 OH· units/h per 105 cells, P < 0.05).
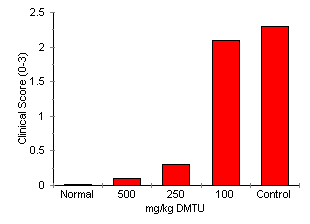
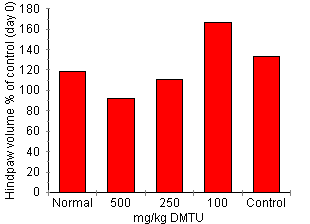
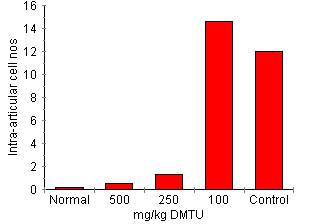
DMTU Effect On Circulating Neutrophil Number Blood (20 DMTU Effect on AA Development Treatment with DMTU significantly attenuated AA in a dose related fashion (Fig. 3). Dimethylthiorea 500 mg/kg completely inhibited joint inflammation. Dimethylthiorea-treated rats had a clinical score of 0 (disease control = 2.3 ± 0.3, P < 0.0001) and did not develop paw swelling (DMTU treated = 91.9 ± 3.3%, disease control = 134.4 ± 8.5%, P < 0.0001). Inflammatory cell accumulation in joints was also significantly reduced (DMTU treated 0.5 ± 0.1 x 105 cells/four joints, disease control 11.76 ± 1.7 x 105 cells/four joints, P < 0.0001). Dimethylthiorea 250 mg/kg also significantly attenuated the clinical score (0.3 ± 0.2, P < 0.05), paw swelling (111.0 ± 5.0% of day 0 volume, P < 0.05) and inflammatory cell accumulation (1.3 ± 0.6 x 105 cells/four joints, P < 0.05): 100 mg/kg DMTU did not significantly affect development of AA. These animals had a clinical score of 2.1 ± 0.6, 1.66.3 ± 17.8% change in paw volume and a cell count of 14.6 ± 7.0 x 105 cells/four joints.
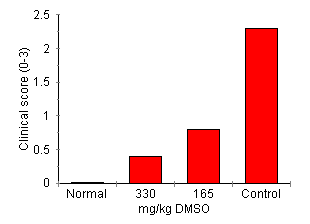
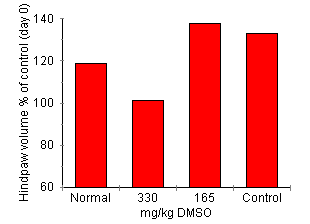
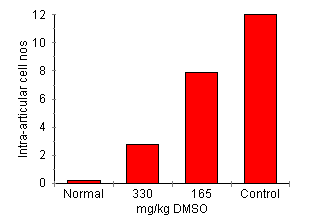
DMSO Effect on AA Development Joint inflammation was attenuated by DMSO treatment in a dose-dependent manner (Fig. 4). Rats treated with 330 mg/kg of DMSO had significant reduction in arthritis indicated by lower clinical score (0.4 ± 0.3, P < 0.01), reduced paw swelling (101.3 ± 7% of day 0, P < 0.01) and attenuation of joint inflammatory cell influx (2.75 ± 1.9 x 105 cells/four joints, P < 0.01) compared with control treated rats (all parameters P < 0.01 cf. disease control). Treatment with 165 mg/kg reduced the clinical severity of arthritis (clinical score of 0.8 ± 0.4, P < 0.05) but failed to significantly reduce paw swelling (paw volume 137.61 ± 8.9% of control) or inflammatory cell accumulation (cell count 7.9 ± 5.6 x 105 cells/four joints.)
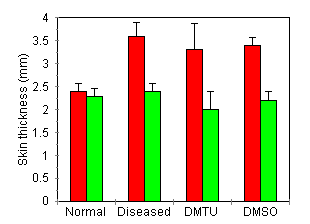
DMTU Effect on Circulating and Peritoneal Neutrophils Rats developing AA showed a significant leucocytosis on day 14 (65 ± 7 x 103 cells/mm3) compared with normal rats (10 ± 0.5 x 103), P< 0.0001). Neutrophils made up 62% of this population with a count of 40 ± 6 x 103 cells/mm3. Treatment with DMTU (500 mg/kg) did not affect the total leucocyte count (64 ± 25 x 103 cells/mm3) or the neutrophil count (40 ± 2 x 103 cells/mm3). Cells obtained from peritoneal lavage were at least 95% neutrophils by nuclear morphology. Peritoneal neutrophil counts from DMTU treated rats (8.1 ± 1.5 x 107 cells/animal) did not significantly differ from counts obtained from untreated rats (9.9 ± 3.2 x 107 cells/animal.) Cutaneous DTH response to disease initiating antigen Maximal doses of DMTU (500 mg/kg) and DMSO (330 mg/kg) did not inhibit cutaneous DTH responses to the disease initiating antigen (Fig. 5). Rats developing AA developed increased skin thickness in response to challenge with MT in oil (3.6 ± 0.3 mm) which was significantly greater than the response to oil alone (2.4 ± 0.1 mm, P < 0.01). Maximal DMTU-treated rats developed cutaneous swelling similar to disease controls (MT in oil 3.3 ± 0.6 mm, oil alone 2.0 ± 0.4 mm, P < 0.01). Similarly changes in skin thickness in DMSO-treated rats was similar to disease controls (MT in oil 3.4 ± 0.1 mm, oil alone 2.2 ± 0.1 mm, P < 0.01). Normal rats did not exhibit any specific Cutaneous response following i.d. challenge with MT (MT in oil 2.4 ± 0.1 mm, oil alone 2.3 ± 0.1mm.)
Adjuvant arthritis in rats exhibits similar clinical and pathological features to human RA. 14 Like RA, AA is destructive, relapsing polyathropathy of distal joints characterized by inflammatory cell infiltration with predominance of neutrophilis in synovial fluid. Adjuvant arthritis is initiated by a T cell response to a mycobacterial wall Ag, which may share epitopes with neoantigens within the joints. 15 The disease can be prevented by T cell depletion and can also be adoptively transferred by T cells. 16, 17 This T cell initiated immune response results in the accumulation of T cells, macrophages, and neutrophils in the joint cavity and peri-articular tissues. 3, 16, 17 The mechanisms by which these potential effect cells cause injury are largely unknown. Neutrophils are the most abundant cell in the synovial fluid of patients with RA and in rats with AA. Their cellular products may thus play a pivotal role in the mediation of joint inflammation. Neutrophils produce a variety of potentially injurious products which may mediate joint injury including cytokines, proteases, OH· and HOCl from ROS and nitric oxide. 6, 18 Enhanced nitric oxide production has been demonstrated in serum and synovial fluid of patients with rheumatoid arthritis and in cultured synovial cells of arthritic rats. 6, 19, 20 These studies together with observations that inhibiting nitric oxide formation attenuates the development of arthritis in rats suggest the nitric oxide may contribute to inflammatory arthritis. 18-20 Hypochlorous acid may also play an important role in joint injury. It is produced from H202 and CI¯ via MPO and can inactivate proteins inhibitors, potentially allowing the unopposed action or proteinases in the joints. It has been suggested that HOCl may predominate over OH· as the major radical produced by the neutrophil oxidative burst implying a minor role for OH· in radical mediated neutrophil induced injury. 6 However, a recent report that a patient with complete functional deficiency of phagocyte MPO but a normal NADPH oridase-dependent ROS developed severe joint injury associated with rheumatoid arthritis, casts doubt on the critical importance of phagocyte MPO-generated HOCl in the development of arthritis. 21 In addition, experimental evidence shows that the HOCl-MPO system is unable to depolymerize synovial fluid hyaluronate even at supraphysiological concentrations, suggesting a poor capacity of the HOCl/MPO system to mediate cartilage destruction. 22 The role of OH· in AA has not been previously reported. Neutrophil OH· production from H2O2 via the Haber-Weiss reaction requires an abundant source of divalent metal ions (e.g. Fe2-). 6 Ferritin is present in higher concentrations in synovial fluid from patients with RA than non-arthritic patients, and contains larger amounts of iron. 23 Superoxide anion can release iron from ferritin. 23 Neutrophil-derived lactoferrin and neutrophil-derived transferring inhibit OH· formation in vitro by binding iron in a non-catalytic form;23 however, these proteins may also be modified by neutrophil proteases to provide cataytic iron complexes. 24 In addition, the capacity of transferrin to bind iron is reduced by the action of ROS. 23 Thus, abundant Fe2- may be available in arthritic joints to allow OH· generation. Neutrophils from synovial fluid of patients with RA have an enhance capacity to generate ROS compared with peripheral blood neutrophils, suggesting local activation of synovial neutrophils. 1 In the current study, unstimulated synovial lavage cells isolated form rats with AA spontaneously produced significantly greater OH· than unstimulated peripheral blood leucocytes from control rats. Hyroxyl radical production of join neutrophils did not increase significantly after further stimulation with PMA. This suggests that neutrophils with arthritic joints are both maximally primed and triggered and produce substantial amounts of OH·. The ROS scavengers, DMTU and DMSO, have been shown to successfully decrease injury in biological systems in which OH· is suspected to contribute to tissue damage. 7, 9-11, 25 In the current in vivo studies, DMTU reduced the clinical severity of arthritis, joint swelling and synovial neutrophil accumulation in dose-dependent manner. At 500 mg/kg, DMTU completely inhibited the clinical signs of AA. Inflammatory cell accumulation in joints was also significantly reduced. Treatment with DMSO also inhibited all three indices of arthritis in a dose-dependent fashion providing further evidence for ROS scavenging as the mechanism of attenuation of injury. Dimethylthiourea and DMSO show a different spectrum of capacities to scavenge other radicals but share the capacity to scavenge OH·. Both DMTU and DMSO react with OH·,7 H2O2,26 and HOCl. 27 Hydrogen peroxide is poorly reactive in aqueous solution and mediates damage after conversion to other oxygen radicals. 6 The role of HOCl in arthritis is yet to be established. Dimethylthiourea is a derivative of thiourea but has substantially less toxicity because of the substitution of its primary amino acid groups which are oxidized to form the toxin in thiourea. 7 As well, DMSO shares the ROS scavenging potential of DMTU, although its reaction leads to formation of other radicals such as methyl radicals after reaction with OH·. 28 The potential for methyl radicals to mediate joint injury is unknown, however as DMSO effectively prevented joint injury in the current studies, this potential pro-inflammatory effect must be minor in AA. The mechanism by which ROS, such as OH·, cause joint injury and inflammatory cell accumulation are uncertain. Reactive oxygen species may injure synovial lining cells, leading to release of chemotactic or pro-inflammatory factors from these cells or from the underlying cartilage. Alternatively, they may alter vascular permeability or expression of cell adhesion moldecules, thus increasing inflammatory cell transmigration. As yet there is no direct evidence to support these hypotheses. Studies by Petrone et al. demonstrate the generation of a potent neutrophil chemotactic factor form ROS exposed plasma, suggesting that ROS may react with plasma proteins to augment neutrophil accumulation at inflammatory sites. 29 Reactive oxygen species may also have a regulatory role in chemokine production as evidenced by the observations that ROS scavengers selectively inhibit interleukin-8 production by LPS-simulated human whole blood. 30 Studies by Weiss, et al.31 and Buckhard et al. 2 also suggest a role for reactive oxidants in the activation of neutropil collagenase. The inflammatory processes in arthritis are mediated by a complex interplay of a number of cellular products. Inhibition of arthritis by depletion of one such product suggests a critical role for it in the development of the disease. It does not exclude a role for other mediators which may also be essential links in the chain of events leading to joint damage. In this study, the demonstration of OH· production by neutrophils harvested form arthritic joints, and the capacity of two different ROS scavengers to inhibit joint injury in vivo, suggest an important role for OH· in the mediation of joint injury in AA.
The advice of Dr. N.W. Boyce and Dr. E.F. Morand in the preparation of this manuscript is gratefully acknowledged. We received grant support from the Lions District 201 VI, Rheumatism and Arthritis Medical Research Foundation and the National Health and Medical Research Council of Australia.
Source: Immunology and Cell Biology (1994) 72, 406-414. DMSO organization wishes to thank the publishers of Immunology and Cell Biology for allowing us to place this article on our World Wide Web site. The publisher retains all copyright. Please contact the publisher for permission to copy any portion of this article. |
|||||||||||
© 2001-2022
All rights reserved |